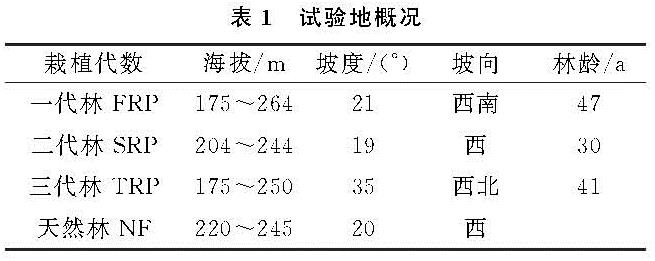
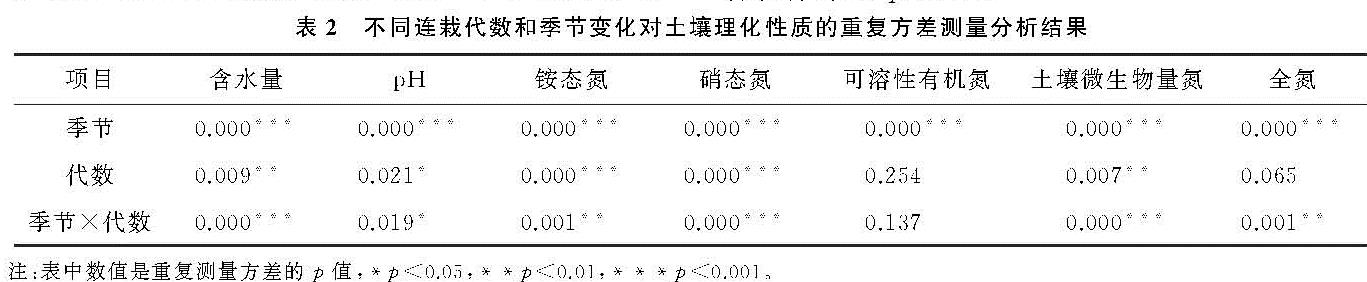
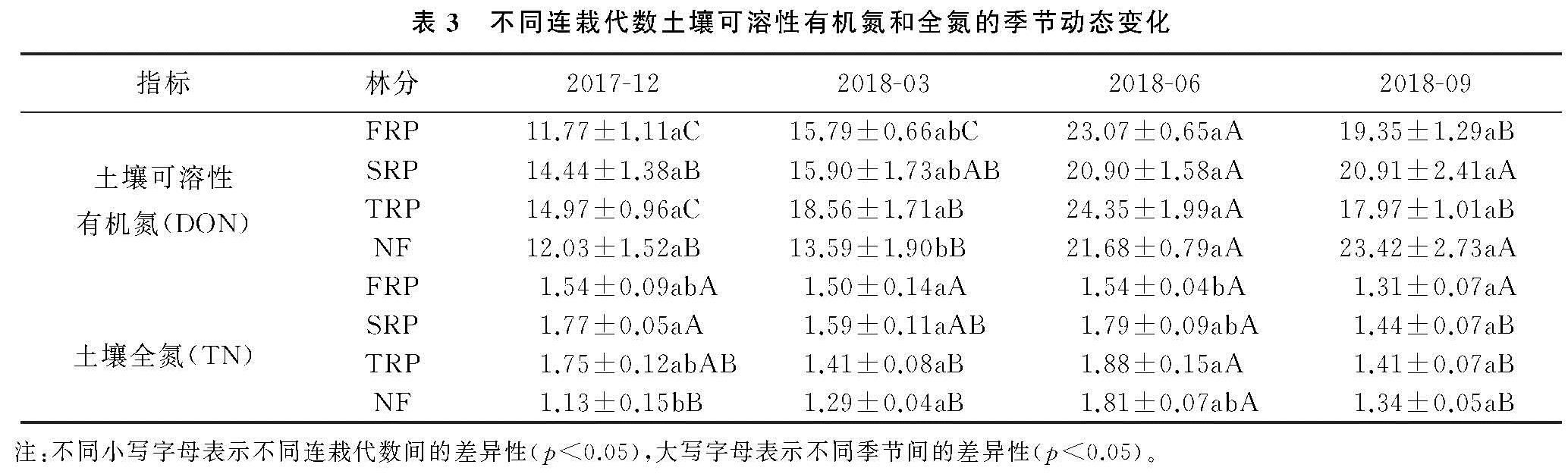
本研究中,不同连栽代数杉木林土壤TN和DON含量均没有显著差异(表2),说明土壤TN没有随着连栽代数的增加而显著减少,这与在广西南亚热带桉树林的研究结果不一致[22],该研究发现1—3代桉树林土壤TN随着栽植代数的增加而减少,且均显著低于前茬马尾松人工林。这可能是由于桉树的轮伐期较短(5~7 a),对养分需求量大,且在采伐时整个树木会被利用而从林地取走,造成林地养分的损失程度大于杉木林。4种林分土壤TN含量最高值均出现在夏季,最低值则主要出现在9月,这与林下植被类型和林内小气候等有关。3月时气温较低,土壤微生物活性和有机氮矿化作用较弱,此时植物开始生长,对有效氮的吸收量逐渐增加,土壤TN的含量的积累逐渐减少; 6月时林木生长旺盛,虽然需要从土壤中吸收较多的氮素来满足其生长需要,但充足的水热条件有利于有机物质的分解,以及凋落物的分解促进了土壤TN的累积; 9月林木生长速度减缓,对氮素的吸收能力仍然较强,水热条件相对夏季较差,导致土壤TN的生物累积能力最低; 进入12月后,虽然土壤微生物活性和有机氮矿化作用最弱,但林木对氮素的吸收能力最低,会促使土壤TN的生物积累较秋季略有增加。森林土壤DON主要来源于凋落物、有机质的淋溶和腐殖质的分解及大气氮沉降[23]。本研究结果显示,格氏栲天然林及连栽杉木人工林4种不同林分土壤DON含量呈明显的季节变化,呈先上升后下降的趋势,其中12月DON含量最低(表3),出现这种趋势可能是因为12月降雨减少且温度低,土壤中微生物的活性低,凋落物的分解速率低,随着降雨量和温度的升高,土壤微生物活动增强,促使土壤动植物体分解,从而使土壤DON含量逐渐增高。
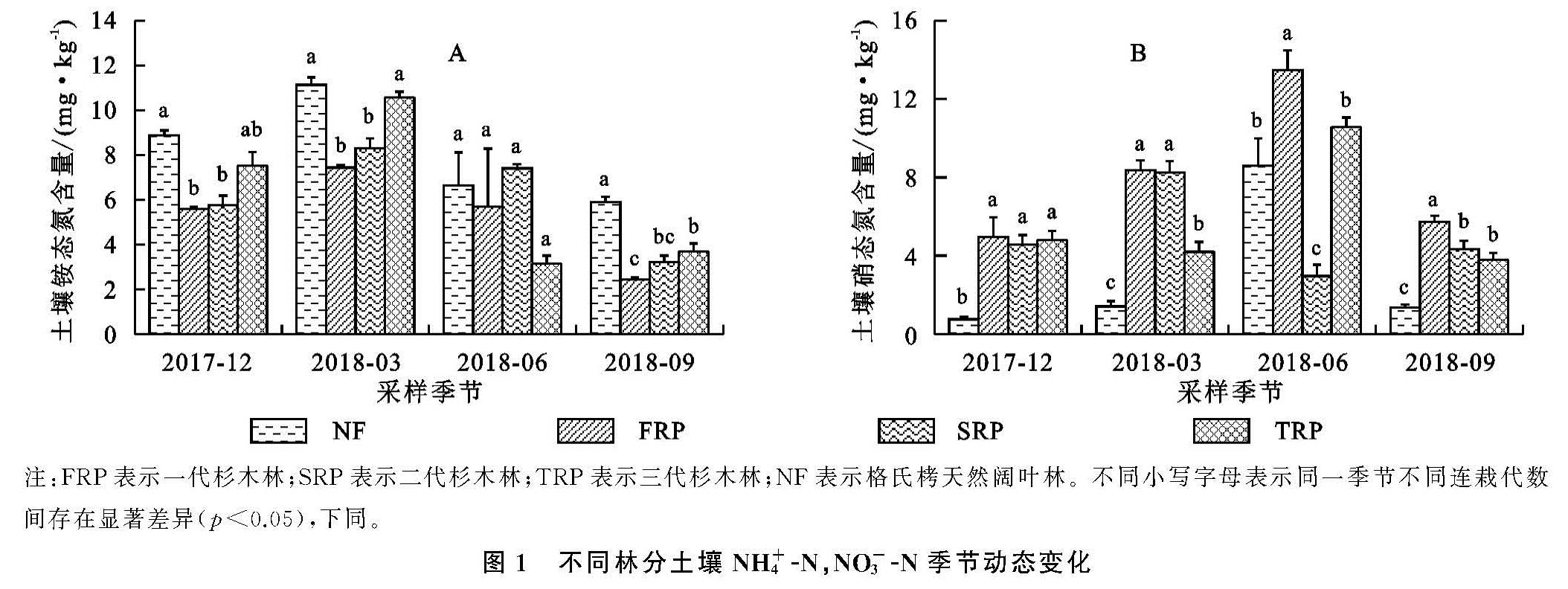
随着连栽代数的增加,杉木人工林土壤NH+4-N总体呈现出增加的趋势(图1A),而NO-3-N则呈现出下降的趋势(图1B),这与Chen等[24]在湖南会同连栽杉木林中发现的结果一致。土壤NH+4-N带正电,更容易被土壤胶体吸附存留于土壤中,而NO-3-N带负电,移动性强,特别是在酸性土壤中更容易流失[25]。在连栽杉木林中土壤氮多以NO-3-N的形式损失[26]。土壤氮有效性很大程度上通过氮矿化过程调控,该过程在微生物参与下将土壤有机氮转化为无机氮。相较于杉木一代林,二代林和三代林土壤NH+4-N升高,NO-3-N下降,可能反映了在土壤氮匮乏条件下系统为了适应环境而采取的保氮机制。随着连栽代数增加,尽管土壤TN含量未见明显下降(表3),但土壤矿化率升高,硝化率降低,从而减少了土壤氮流失的风险,这也与Chen等[27]对连栽杉木林中土壤净矿化率和硝化率的测定结果一致。同样,在江苏北部杨树人工林的研究也发现,二代四年生杨树林土壤NO-3-N含量(7.67 mg/kg)显著低于一代四年生林地(30.93 mg/kg),这是由于二代杨树林土壤硝化作用显著减弱导致土壤供NO-3-N的能力减弱,需要注意的是,该研究中测定的净硝化作用,要想深入了解连栽对土壤硝化能力的影响,还需借助氮同位素示踪技术测定土壤总硝化速率、微生物对NO-3-N的同化吸收速率、以及反硝化过程中气态氮的损失速率[28]。有报道表明,随着连栽代数增加,杉木林土壤中氨氧化细菌的数量和种类均有所下降[29],这可能是造成土壤硝化作用减弱和NO-3-N含量降低的原因。未来研究可结合参与硝化与反硝化功能微生物的数量与不同形态氮的关系深入探讨连栽杉木林土壤氮的转化和去向。值得注意的是,无论在哪个季节,杉木人工林的土壤NH+4-N含量普遍低于天然林,而NO-3-N含量则高于杉木人工林,说明天然林转化为杉木人工林会使土壤氮淋失的风险增高。尤其本样地位于亚热带季风气候区,夏季降雨充沛,更不利于土壤速效氮的截留与保持。
土壤矿质氮的动态变化在很大程度上依赖于土壤微生物对土壤有机质的分解速率和微生物固持、气态损失和淋溶等多种过程的相互竞争[11,17]。本研究中,不同林分土壤中NH+4-N和NO-3-N含量呈明显的季节变化,主要受局地小气候影响。土壤N素转化过程主要靠微生物参与,而微生物对土壤含水量和温度反应非常敏感[30]。其他地区不同林分的研究结果则与本研究样地有所不同,如四川都江堰地区桢楠林、杉木林以及常绿阔叶研究显示土壤NH+4-N在冬季(12月份)达到最大值[31],重庆缙云山地区4种典型林分表层土壤(0—10 cm)NH+4-N和NO-3-N的最大值均出现在秋季[32]。可见由于研究地区以及林分类型不同,土壤的理化性质、生物特性以及气候都存在差异,从而造成研究结果不同。报道也显示,NH+4-N和NO-3-N含量有明显的季节变化性和空间变异性[33]。
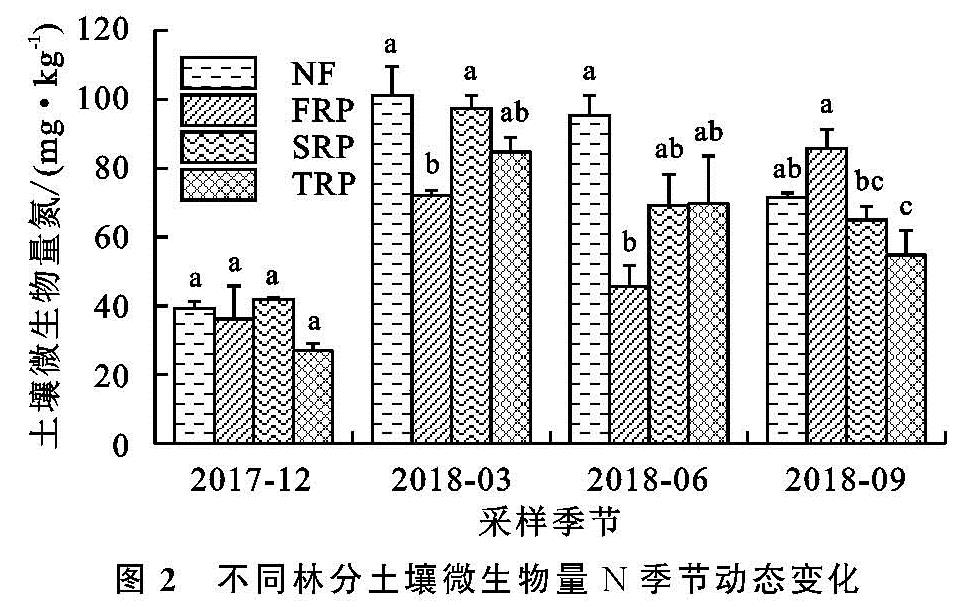
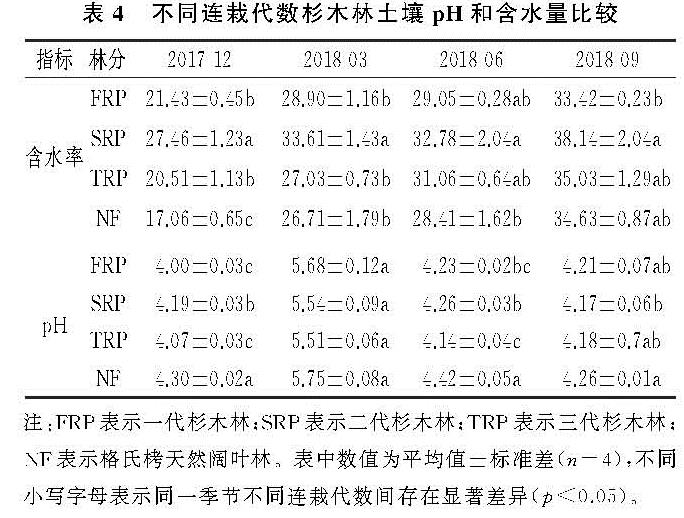
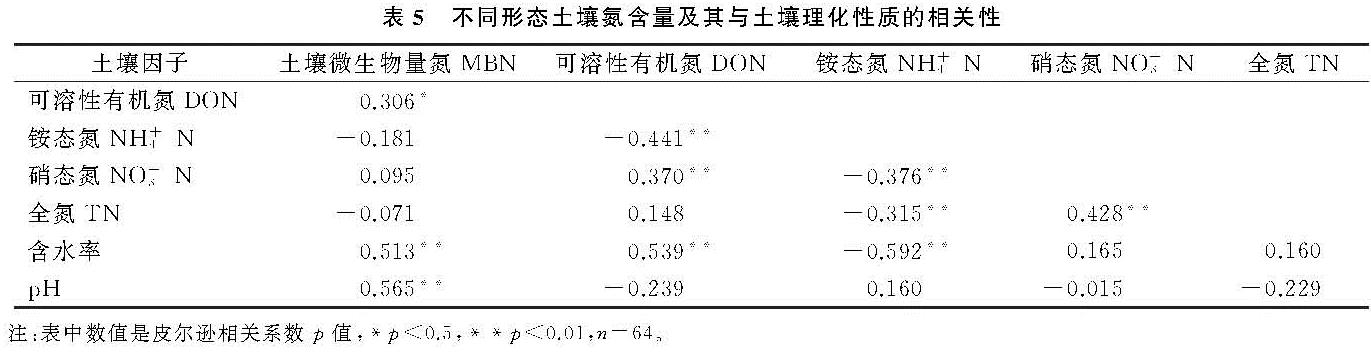
土壤MBN是土壤有机氮中最活跃的组分,也是土壤氮循环和转化过程中的重要参数[34]。土壤微生物量的季节性动态变化是一个复杂的过程,在同一生态系统,即使气候条件相同,不同植被下土壤微生物量的季节变化也不相同[35-36]。土壤微生物量的变化与温度、湿度、土壤理化性质等环境因素有关,且土壤微生物因季节变化也会有差异,微生物的群落结构和功能也随之变化,从而使微生物量因季节变化而变化[37]。本研究结果表明,土壤MBN与土壤水分呈显著正相关关系(表5),表明该区域土壤含水量是影响土壤MBN的重要因子之一,这与Chen等[27]研究发现一致。此外,在连栽杉木人工林内,相关分析表明,土壤MBN与土壤pH呈显著正相关(表5),这表明土壤酸碱性也是影响土壤微生物量的一个重要因素。本研究中4种林分的土壤MBN含量具有明显的季节变化(表2),二代林、三代林和天然林均呈现出先增加后减少的趋势(图2),其中6月、9月为植物的生长期,MBN含量较低,这与大多数研究发现的土壤MBN有季节变化的结果一致[38-39]。植物在生长期对土壤养分的需求量大,限制了土壤微生物对养分的可利用性,因此生长期土壤微生物生物量氮较低。本研究中土壤MBN都是在春季(3月)最高(图2),说明春季在土壤温度、湿度还比较适宜的时候,植物生长缓慢,植物对N的竞争减少,因此导致土壤MBN增加。4种林分中一代林土壤MBN含量的季节动态与其他3种林分不同,从12月到3月土壤微生物量N含量增加,这可能是因为在冬季与春季交替的过程中,水分增多、温度升高,微生物活动频繁,土壤MBN快速增加; 到6月温度持续升高,微生物活动强,资源消耗快,由于竞争导致MBN含量下降; 9月以后植物生长速度变慢,温度降低、降雨减少,且凋落物由上而下的转移,营养物质的输入导致土壤MBN含量开始增加。土壤微生物组成结构不同,且不同的微生物种类会对季节变化产生一定影响,从而使土壤MBN含量有明显的季节变化。