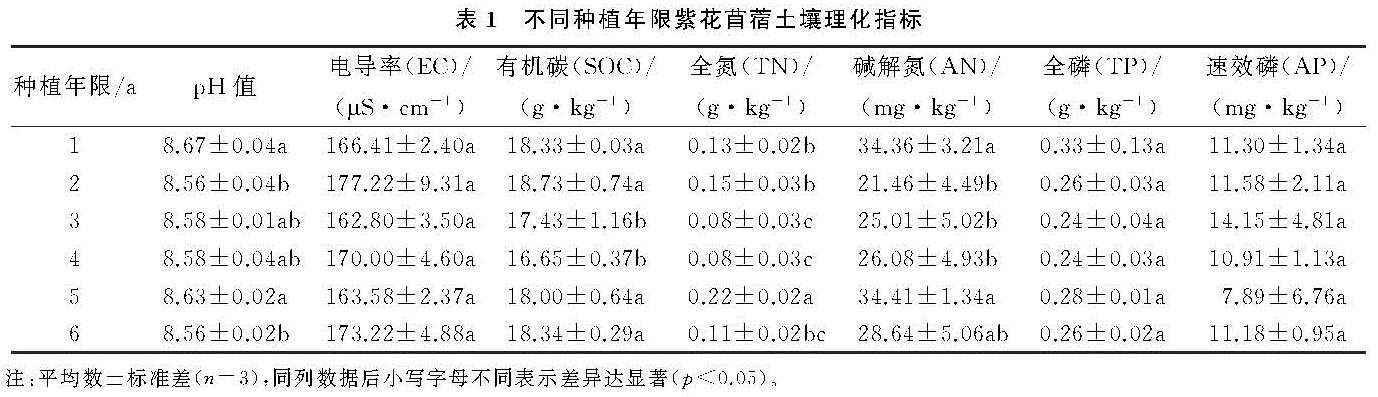
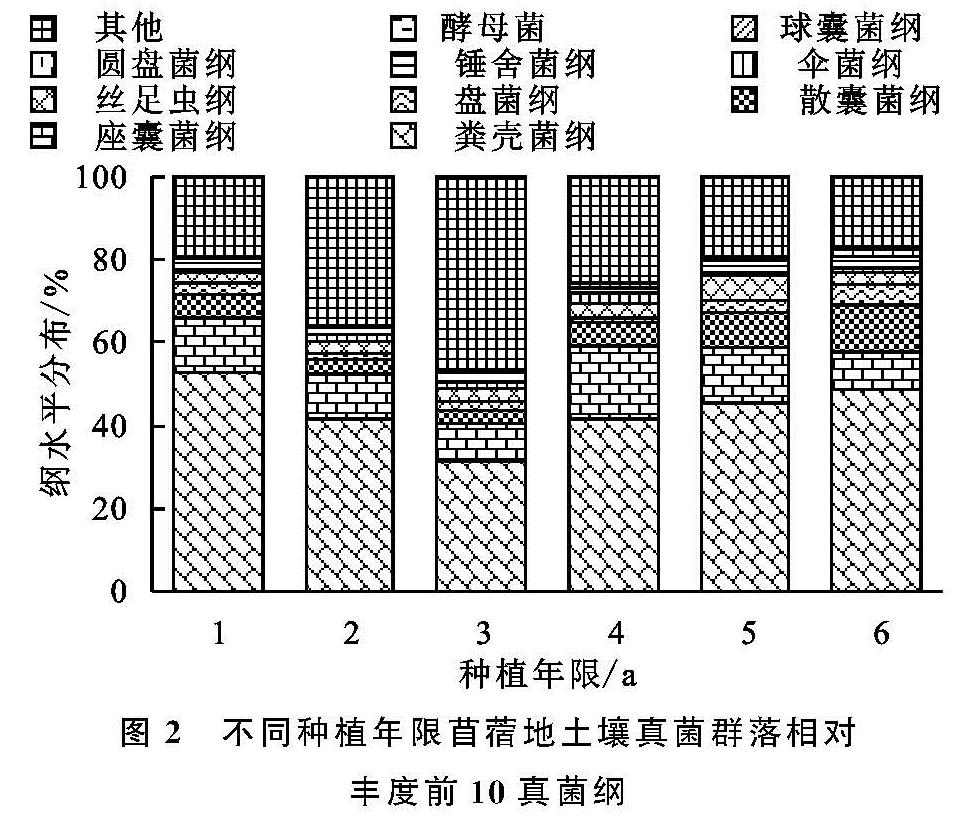
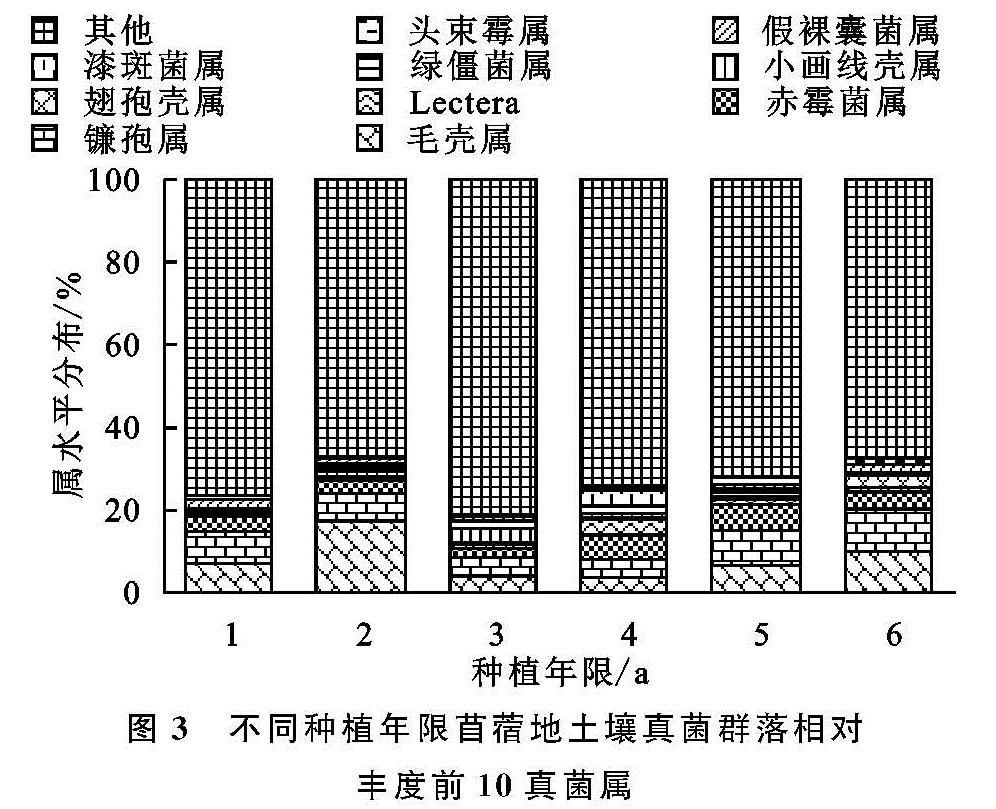
土壤真菌数量大,种类多,是检测土壤质量变化的重要指标,在评价土壤生态系统等功能方面具有重要作用[23]。本研究通过对不同种植年限苜蓿地土壤在纲水平和属水平下真菌群落组成中丰度占比前10的菌类分析发现,粪壳菌纲、座囊菌纲、散囊菌纲、盘菌纲、丝足虫纲为优势菌纲,毛壳属、镰孢属、赤霉菌属、Lectera、小画线壳属为优势菌属。其中粪壳菌纲占总纲类真菌的44%,而且随着种植年限的增加,没有大的波动,粪壳菌是一种寄生在动物粪便上的真菌,因此粪壳菌成为最具优势菌的原因可能是农田管理过程中大量农家肥施入的结果,其他优势菌类随着种植年限的增加,表现为先减小后增大的变化趋势,这与土壤理化指标变化基本保持一致。在属水平下,土壤真菌群落由于受pH值,AN和SOC等理化因子的影响,随着种植年限的增加,各菌属变化不再呈现规律性变化。相关的研究显示,镰刀属、毛壳属和木霉属在土壤中是常见的纤维素分解者[24],而本研究发现毛壳属为6个样本中最具优势的真菌,占总菌属的8%,因此毛壳属可能是影响苜蓿地土壤碳循环的主要真菌菌属[25]。
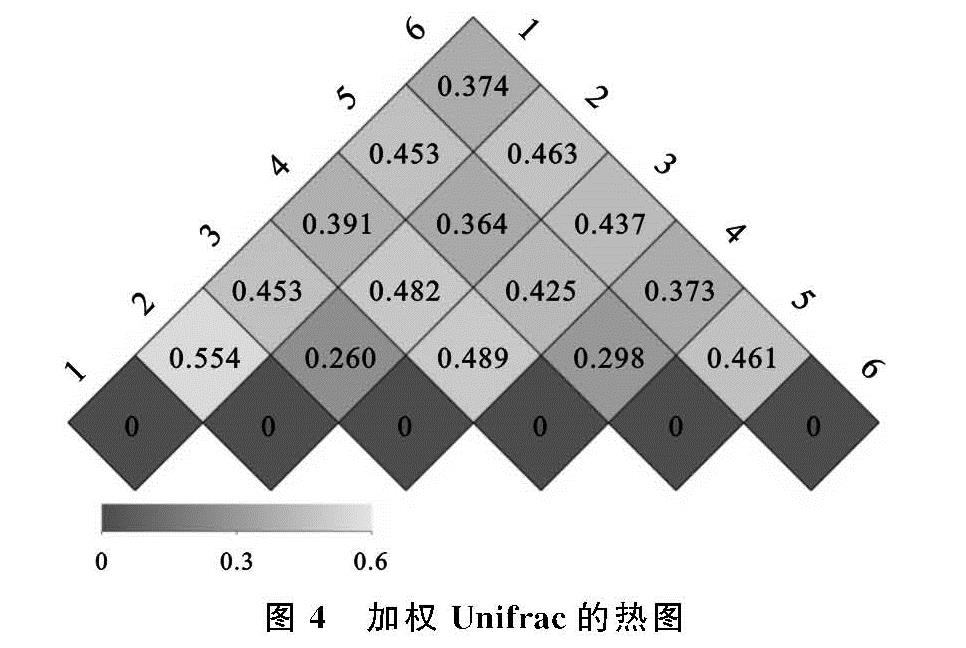
土壤微生物群落多样性通常用多样性指数的大小来分析,多样性指数越高则微生物群落多样性越高[26]。本试验通过对指示群落结构变化的Alpha多样性指数进行组间差异性分析,最终研究结果表明,Simpson指数不受种植时间的影响,而Shannon指数、Chao1指数和ACE指数表现出相同的变化规律,即均在种植2 a时达到最小,5 a时达到最大,且差异达显著水平,因此连种苜蓿会使土壤中真菌菌群的多样性处于一个较高的水平,这与Wal等[27]发现休耕地土壤真菌多样性指数也处于较高水平的研究结果保持一致,毕明丽[28]、孙倩[29]等研究结果也支持这一结论。Beta多样性分析结果显示,随着种植年限的增加,土壤真菌多样性变化表现为由变幅较大到逐渐趋于平稳的趋势,各种植年限间相异系数可反映这一变化规律。
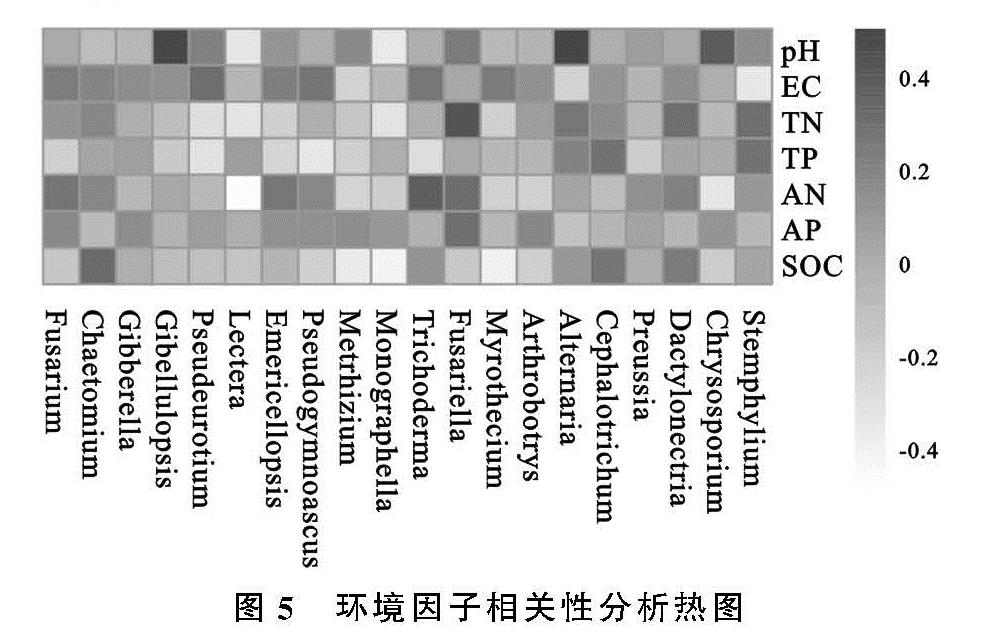
已有研究表明,土壤真菌多样性和群落组成受多种因素影响,土壤环境是其中一个重要因素[30-31],因此对属水平下真菌群落组成与土壤环境因子进行Spearman相关性分析,有助于探索影响不同种植年限苜蓿地土壤真菌群落结构及多样性的主要影响因子。结果表明土壤pH值,AN和SOC与一部分真菌间存在显著或极显著相关性,EC、TP和TN与个别真菌存在显著相关性,AP与所有真菌不具有相关性。Liu等[32]研究发现,速效磷对土壤真菌群落结构具有较大影响,且施磷肥会提高土壤真菌多样性[33],而本研究中,速效磷变化范围为7.89~14.15,随着种植年限的增加未发生显著性变化,因此未能体现影响程度。Beare等[34]提出免耕少耕能增加真菌菌群的多样性,增加不稳定碳的固定积累,进而影响土壤有机碳含量,这与本研究Spearman相关性分析结果一致。本研究中pH值随着种植年限的变化第1年与第6年存在显著性变化,且pH值与真菌属水平下存在显著或极显著相关性,这与韩世忠等[35]研究发现土壤pH值会对真菌群落结构组成造成影响的结果一致。