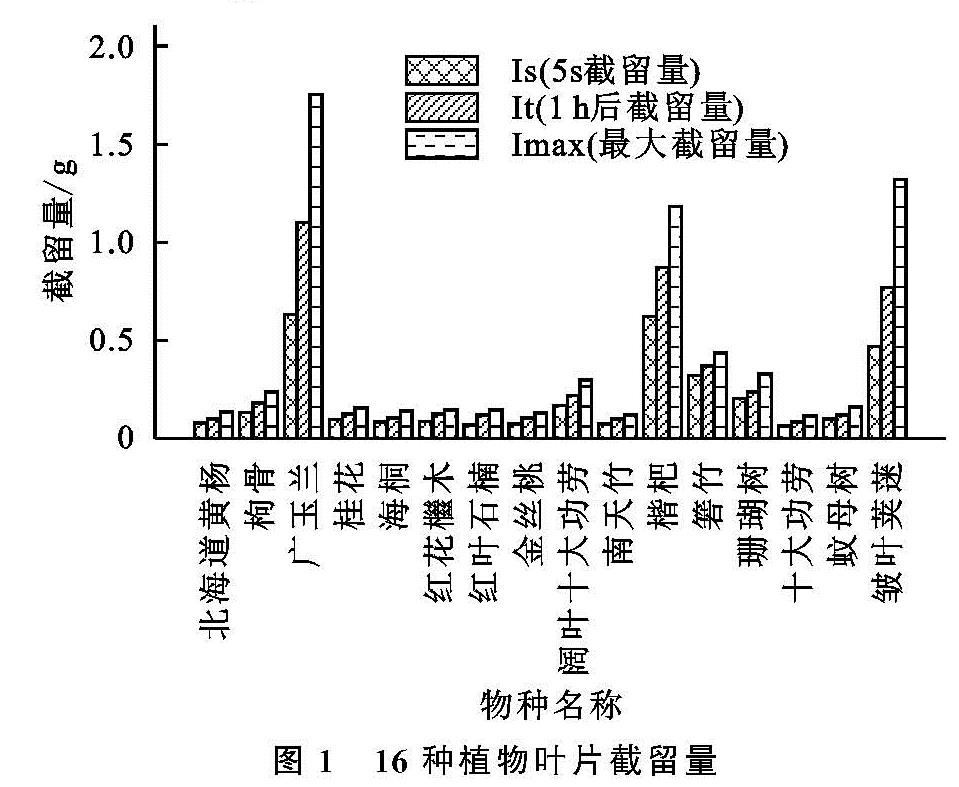
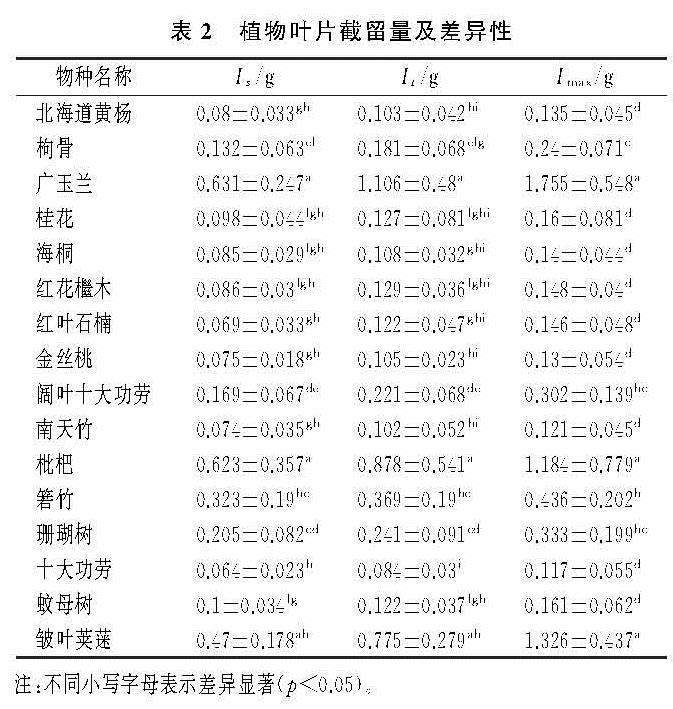
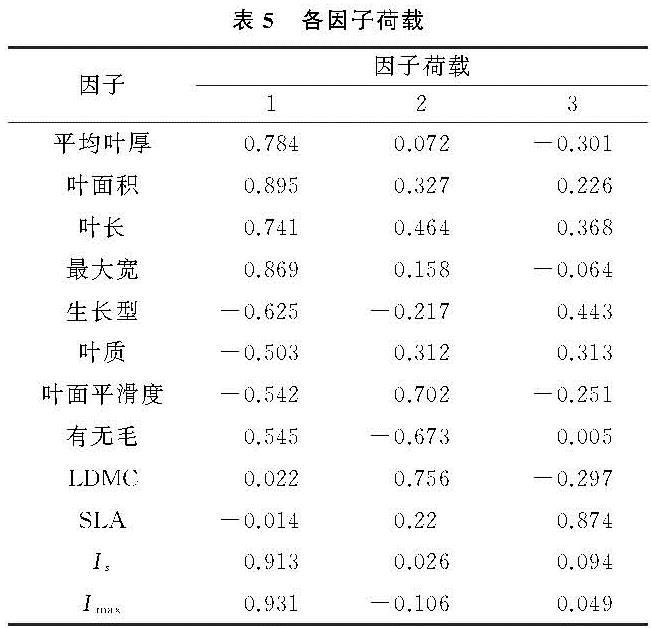
(1)本研究中对叶片截留能力影响最大的叶片性状因子有叶面积、叶长、叶宽、叶厚、SLA及LDMC。有研究证明叶面积大小[10]、叶长[14]、叶宽[21]对叶片截留能力的影响显著,本研究中广玉兰、枇杷、箬竹、皱叶荚蒾的单叶截留量较大,与已有研究的结果相同。叶片形态越大,则与水分的接触面越大,因此截留量越大。有关叶厚对截留能力影响的研究较少。叶片纸质的箬竹虽叶片厚度较小但单叶截留量较大,这可能是因为箬竹的单叶面积在16种植物中仅次于广玉兰,属于叶面积较大植物。除箬竹外,总体而言,本研究中叶片厚度与截留能力为显著正相关,符合植物通过增加叶厚来适应干旱强光的环境,进而达到降低蒸腾作用,减少组织内部水分散失,提高水分利用率的现象[22]。
除本研究中PCA筛选出的主要叶片形态因子外,叶毛在前人的结论中被认为也是影响叶片截留持水量的重要因子[13-14]。不同的绒毛类型影响着水滴在叶表皮的存在形式,进而使叶片的湿润性也有所差异[23]。仅以有无来评断叶毛对截留量影响的局限性,以及被试物种的差异可能是本研究与他人研究结果不同的原因所在。
(2)植物叶片功能因子与水分截留的耦合作用,反映了植物适应环境条件的自我调控机制。水分作为光合作用的重要影响因素之一,不仅直接参与光合作用过程,更是通过影响叶绿素合成等间接影响叶片的光合能力[24]。叶片含水量降低,则光合速率增大,水分利用率增大[25]。植物叶片光合作用能力直接影响干物质积累,光合作用越强,LDMC越小,植物叶片的组织密度较低,减小了水分向表面扩散的距离及阻力[26],内部水分散失的增大增加了植物对水分的需求,叶片截留持水能力增加,因而LDMC与Imax间呈负相关。就SLA而言,代表着一定干物质投资所展开的捕光表面积数量,是反映植物相对生长速率的因子[27],这可能是本研究中SLA与截留量相关关系较弱的原因。叶片内部生物量分配是对环境的响应。SLA较高则植物单位生物量的面积较大,叶片吸收水分的表面积较大,因此有利于叶片吸水[20]。SLA较低的植物为降低蒸腾作用,避免过度失水而形成面积较小、较厚或组织密度较大的叶片[28]。因此,为验证SLA与叶片截留能力的作用,进一步探究SLA与被试16种植物的单叶截留率以及平均叶厚的关系,结果显示SLA与单叶截留率呈明显的正相关,与平均叶厚呈明显的负相关,符合较小的SLA可以降低内部水分丧失,提高水分利用率的现象。虽然高SLA的植物叶片对于雨水的截留能力较强,但高SLA的植物生命周期相对较短且对不良环境的适应性更差[29]。这可能是本研究中高SLA树种(皱叶荚蒾、珊瑚树)的单叶截留量反而小于中等SLA树种(广玉兰、箬竹、枇杷)的单叶截留量的原因。
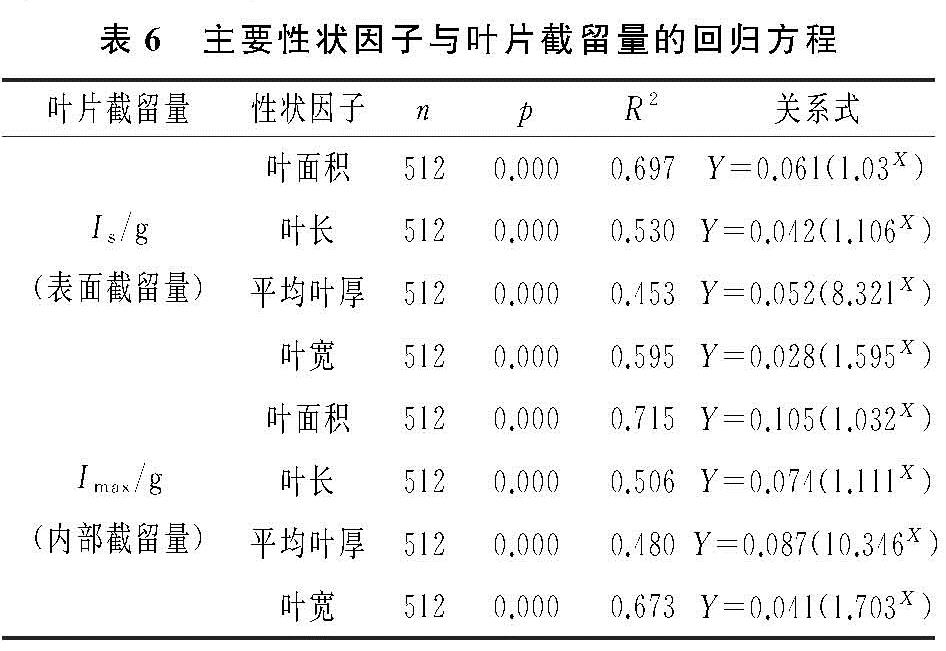
(3)叶片对雨水截留的利用表现为两方面。其一为叶片利用表面润湿性,使水分在叶表面铺展,即叶片表面截留水分。再通过蒸腾蒸发作用使水分返回生态系统,从而实现对水分的再分配,达到增加湿度、促进大气水循环的生态作用。其二是叶片表面的微形态结构及其内部的亲水化合物可以帮助叶片吸收水分,不仅可以截留水分,也可以改变叶片本身的水势,提高叶片的含水率[30]。再通过茎叶间的水势差,使多余水分由叶片进入树种、茎干甚至根部土壤,实现水分从大气到植物根际的逆向运移过程。叶片的内部截留量对植株整体的生理过程及环境适应性产生影响[9]。基于此,本研究探究了叶片性状因子与表面截留量、内部截留量的关系,综合二者来表现叶片性状因子与叶片截留能力的关系。
(4)通过对关中地区16种常绿植物叶片性状及截留能力的定性定量分析,得出叶片形态较大、LDMC较小、SLA中等水平的植物截留能力最佳,即广玉兰、箬竹、枇杷等。但本研究中截留量测定的时间跨度较大,因此未能准确反映植物叶片截留速率变化的具体时刻差异。后续研究中可细化时间间隔,并进行一年四季的测定,以得到较为完整的叶片截留变化态势。此外,本研究探讨的叶片形态特征仅为宏观形态,可以从叶片微观结构层面深入探究叶片内部不同结构类型与水分截留的具体关系。