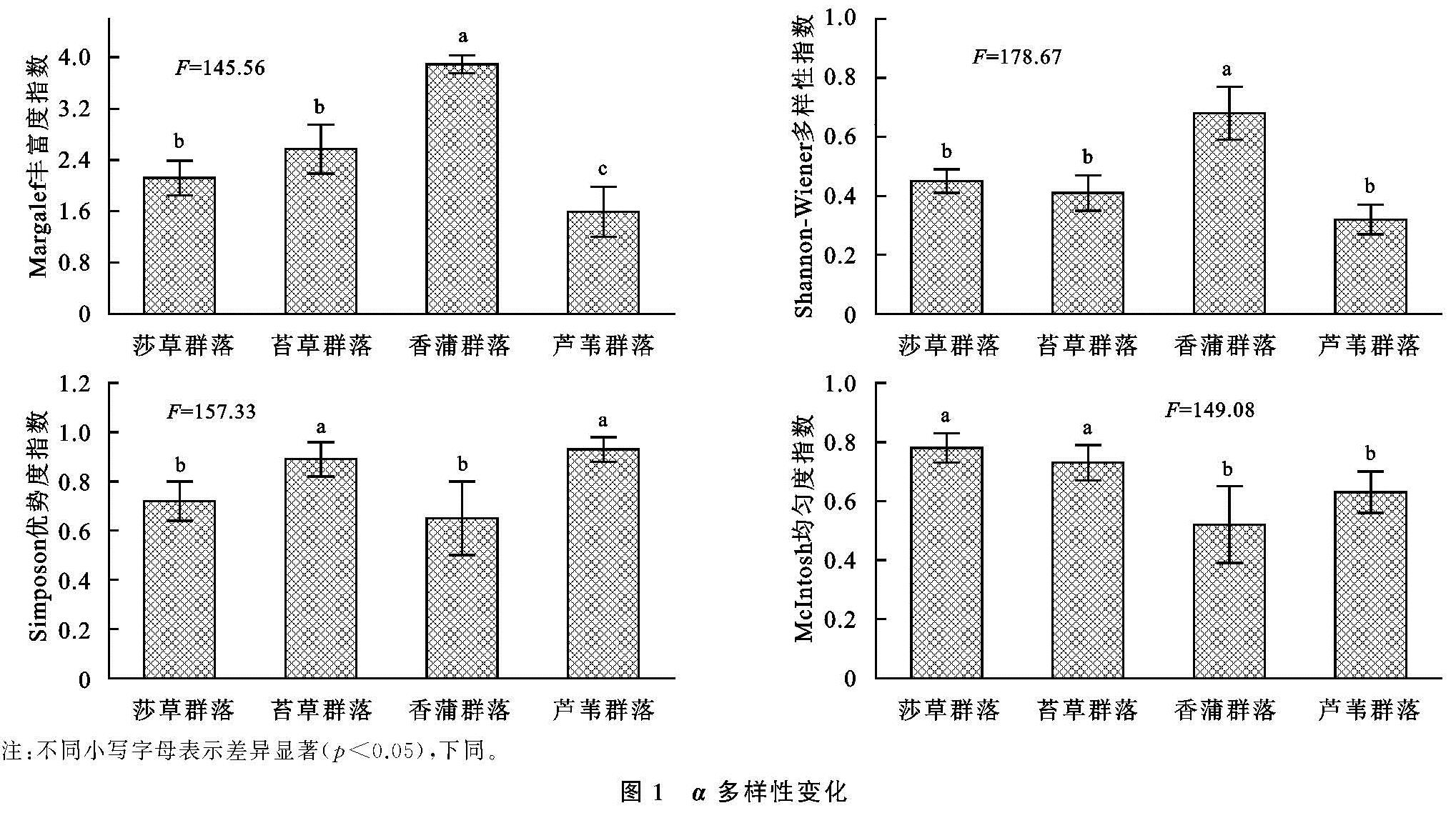
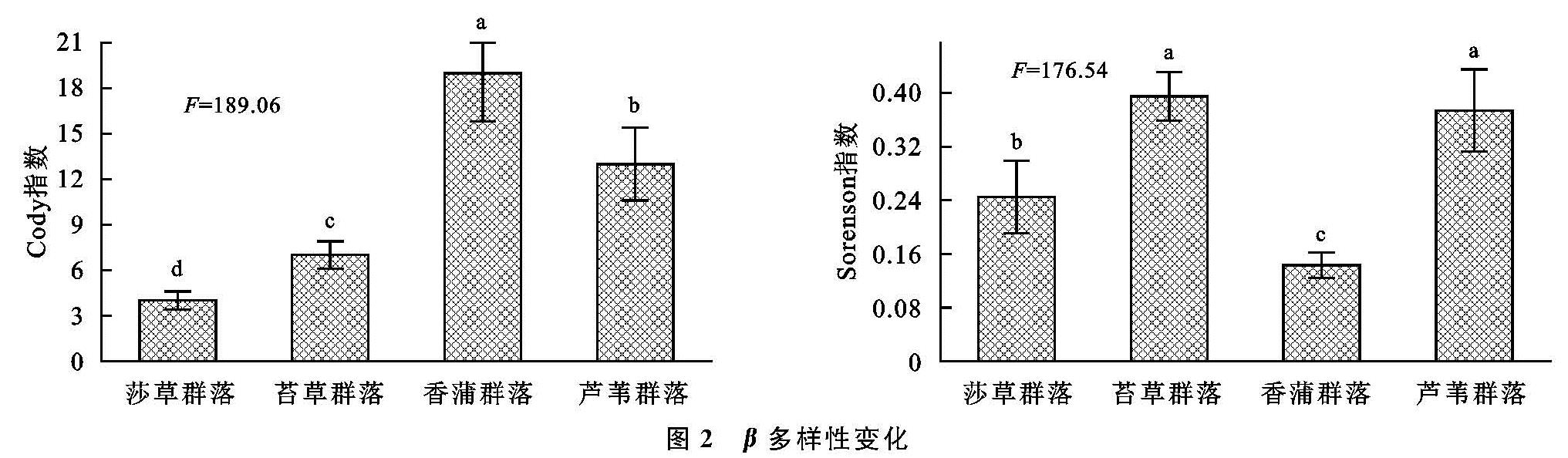
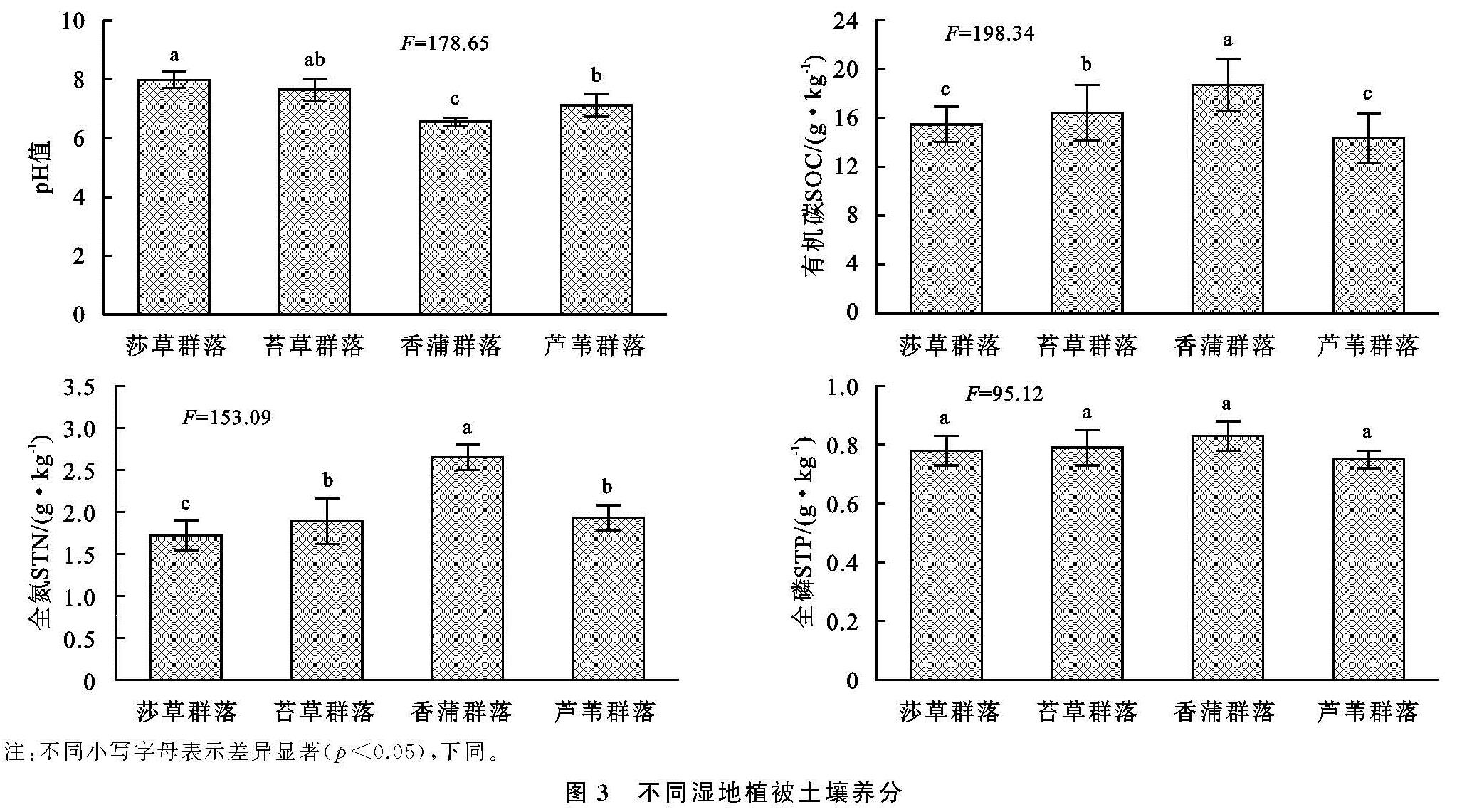
对于湿地生态而言,其中多种生态子系统充斥其中,其影响因素呈现多样性、复杂性并行的特点,尤为关键的是土壤及微生物等,受制于微生物分布数量和群落特点,其对碳源的利用水平也呈现较为突出的差异[19],为了探究这一差异,通常使用丰度等多样性指数加以研究。对于湿地植被群落分布而言,其影响因素不仅呈现突出的多样性,并在此基础上凸显复杂性,其中尤为突出的是微生物以及酶,在微生物的普遍参与下[20],大量的有机质能够得以有效降解,而在酶这一关键角色的参与作用下,形成高效的养分转化,进而促进湿地植被多样性分布[21]。通过连续5 a的观测分析不难发现,立足于α多样性的角度来说,在Shannon,Margalef丰度方面,虽然植被具有明显的不同,导致其出现较为突出的差异,但是整体而言,其明显呈现较为接近的变化特点,即均在年份不断增加的情况下呈现上升态势,且最高的是香蒲,其次是苔草,而芦苇最低; 对于Simpson,McIntosh均匀度方面,则与丰度呈现较大的不同,其具有最高指数的是芦苇,其次苔草,而香蒲最低; 对于香蒲而言,由于其分布于独特的绿洲边缘,其生长环境更为独特、复杂,因此其多样性分布呈现明显的不同,具有更高水平的α多样性。立足于β多样性这一角度,在时间不断增加的情况下,Cody指数呈现较为突出的上升态势,且最高的是香蒲,其次是苔草,而芦苇最低; 对于Sorenson指数来讲恰好相反。综合来看,对于闽江河口湿地来讲,其植被多样性分布呈现整体的上升态势,尤其是芦苇群落,具有较稳定的生态群落分布; 从Sorenson指数的角度来讲,该指数的下降说明物种不断增多,这主要受制于自然环境及人为因素。对于土壤有机碳及全氮而言,虽然植被群落分布出现大幅差异,但是其分异趋势相差并不明显,总有机碳含量对于全氮施加重要影响,有机碳含量直接受制于有机质分解和利用状况,这将受到腐殖质及微生物分解的制约,对于芦苇而言,其具有较为发达的根系,能够有效利用土壤有机碳,从而形成较高水平的有机碳含量。
对于氮而言,其一方面来源与枯枝落叶等腐殖质,另一方面来自于生物固氮; 对于氮输出而言,其一方面是植被的吸收利用所致,另一方面是硝化及反硝化作用下导致氮输出[19]。对于土壤pH来讲,不同的植被形成了不同的pH值,其值最高的是莎草,其次是苔草,而香蒲最低; 对于有机碳来讲,含量最高的属于香蒲群落,其次是苔草,而含量最低的是芦苇,而全氮含量刚好与之相反。对于全磷含量而言,其基本相差不大。对于香蒲而言,土壤具有更好的含水量,长时间的水质影响之下,其土壤表层并不厚,且其有机质含量并不高,因此其在氮素利用方面并不强。
表2 不同植物群落土壤养分、微生物多样性与植被多样性相关性
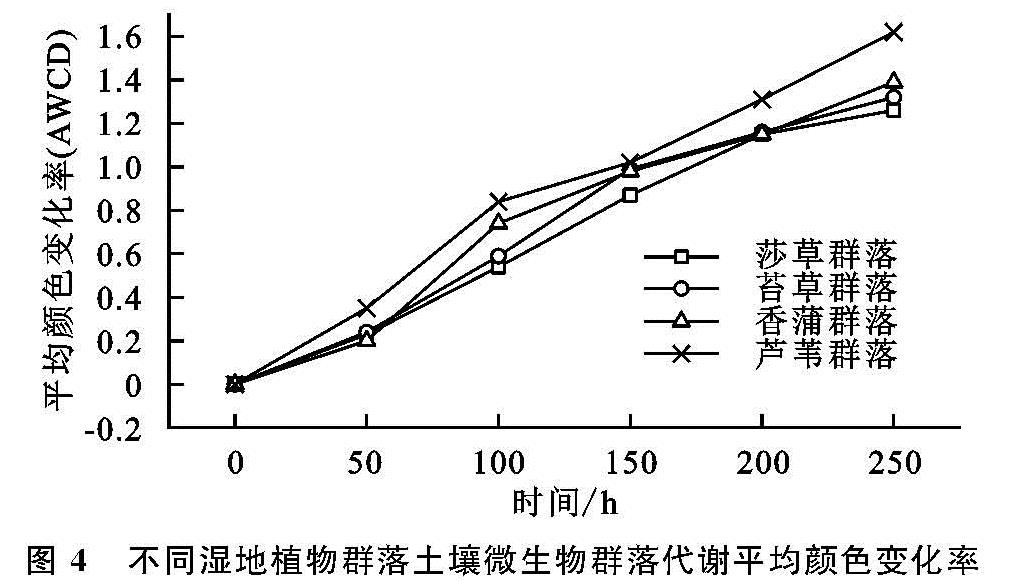
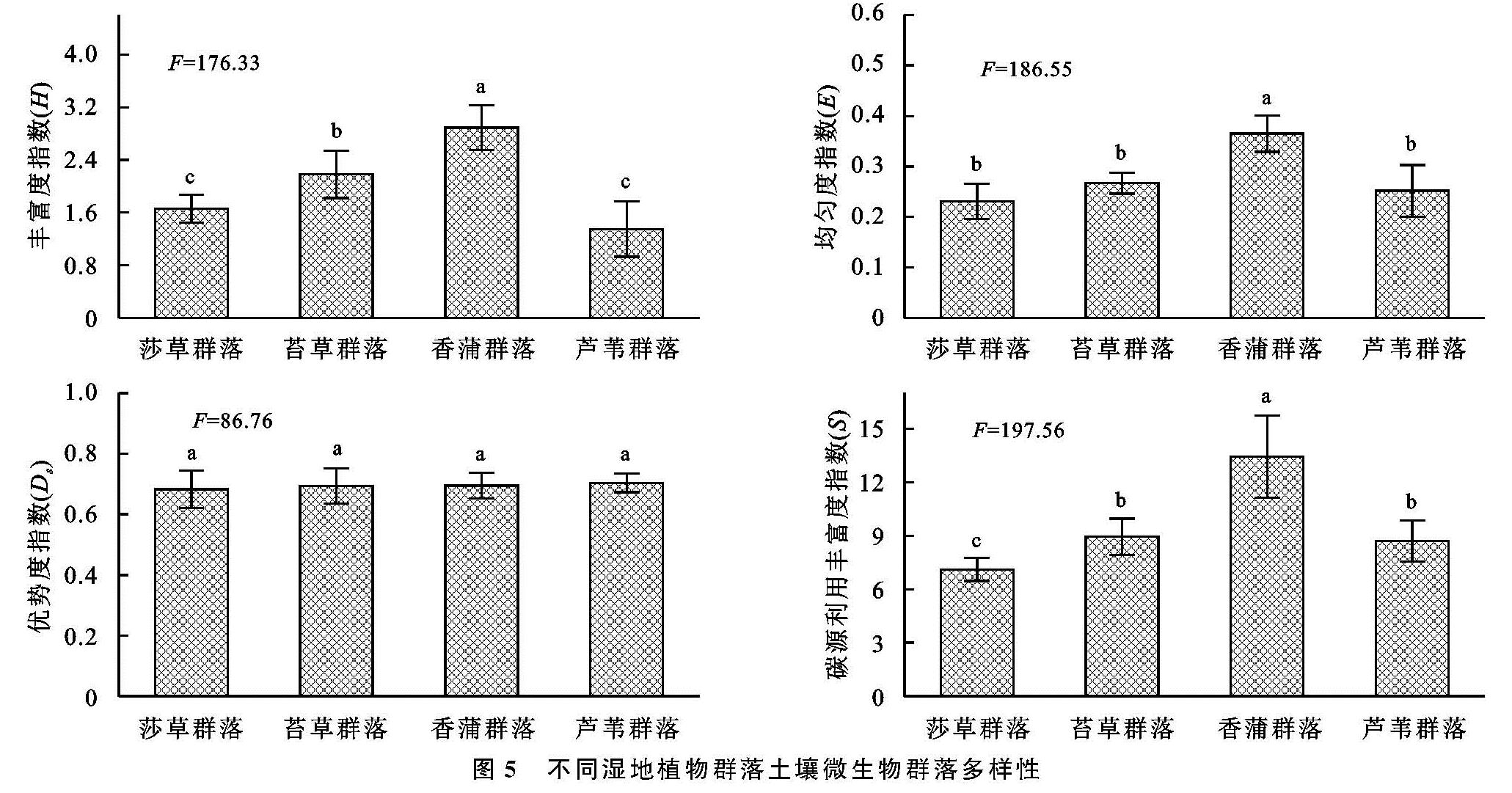
对于湿地植被群落分布而言,其影响因素不仅呈现突出的多样性,并在此基础上凸显复杂性,其中尤为突出的是微生物以及酶,在微生物的普遍参与下,大量的有机质能够得以有效降解,而在酶这一关键角色的参与作用下,形成高效的养分转化,进而促进湿地植被多样性分布。为了对微生物的碳源利用状况进行多角度分析,本研究在Biolog-Eco板的参与下进行有效的展开分析,具体来讲主要借助于AWCD加以衡量,该值越高说明其具有更高水平的碳源利用能力。通过连续5 a的湿地观测分析得知,受不同植被分布影响,其微生物分布及活性也并不相同,无论是丰度H、均匀度E,还是碳源利用丰度S来说,虽然植被具有明显的不同,导致其出现较为突出的差异,但是整体而言,其明显呈现较为接近的变化特点,即指数最高的是芦苇[21],其次是香蒲,而莎草最低; 而对于优势度指数Ds来讲,不同植被间的差异并未达到显著水平[22]。
为了探究闽江河口湿地微生物与植被之间的复杂关系,本研究开展了相关分析,从多样性的角度来看,二者具有紧密关系,这说明微生物新陈代谢等活动能够显著地制约着植被分布多样性,反过来植被状况也能够对微生物状况产生制约,二者相辅相成。为了探究不同影响因素的交互作用,本研究开展了冗余分析,对于河口水质而言,其不仅受到植被及微生物的单个因素制约,同时受制于二者的交互作用影响,且交互作用产生的影响更强。综合来讲,多种环境因子及微生物等制约着闽江河口湿地植被分布,土壤环境的保持离不开微生物,二者是相辅相成的,为了有效保护闽江河口湿地,一方面需要注重植被保护,另一方面需要充分利用微生物和土壤质量的协调。