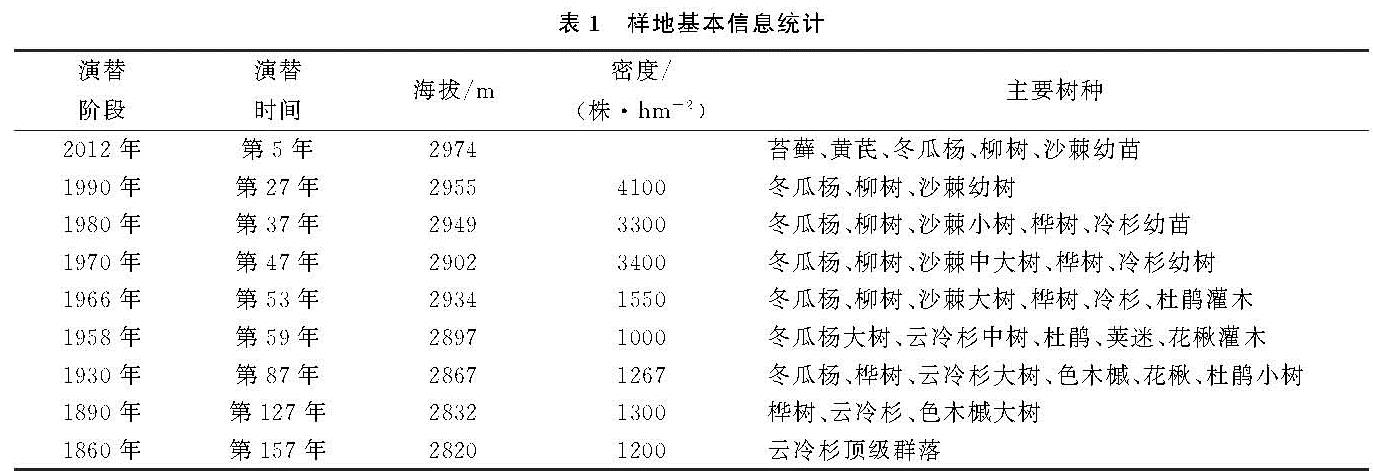
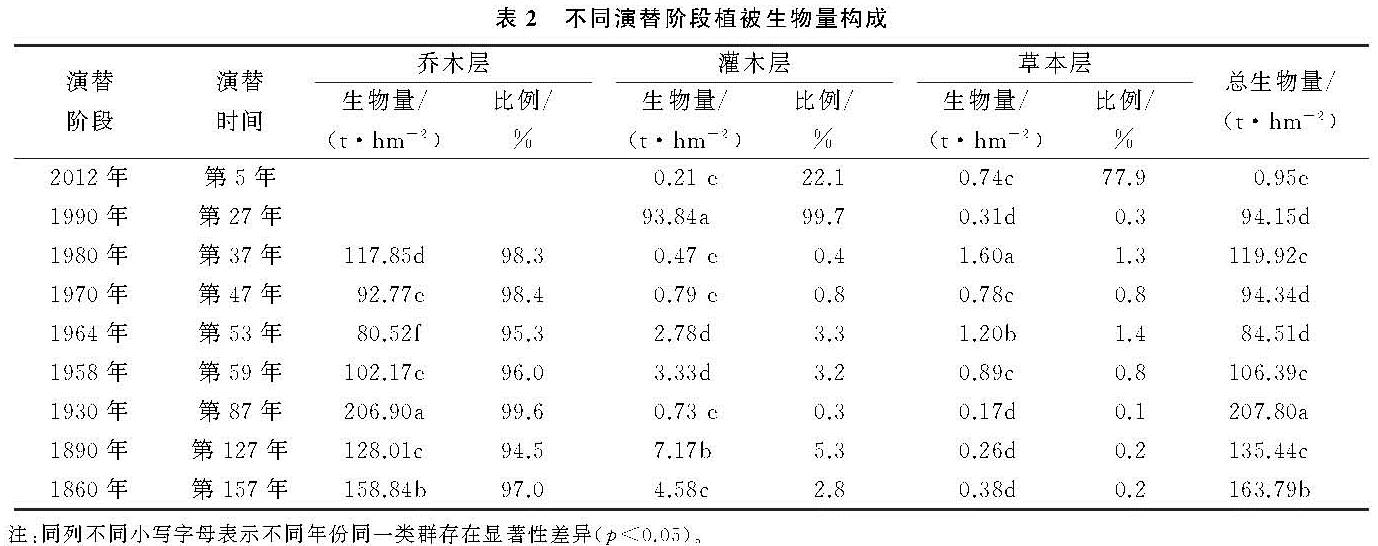
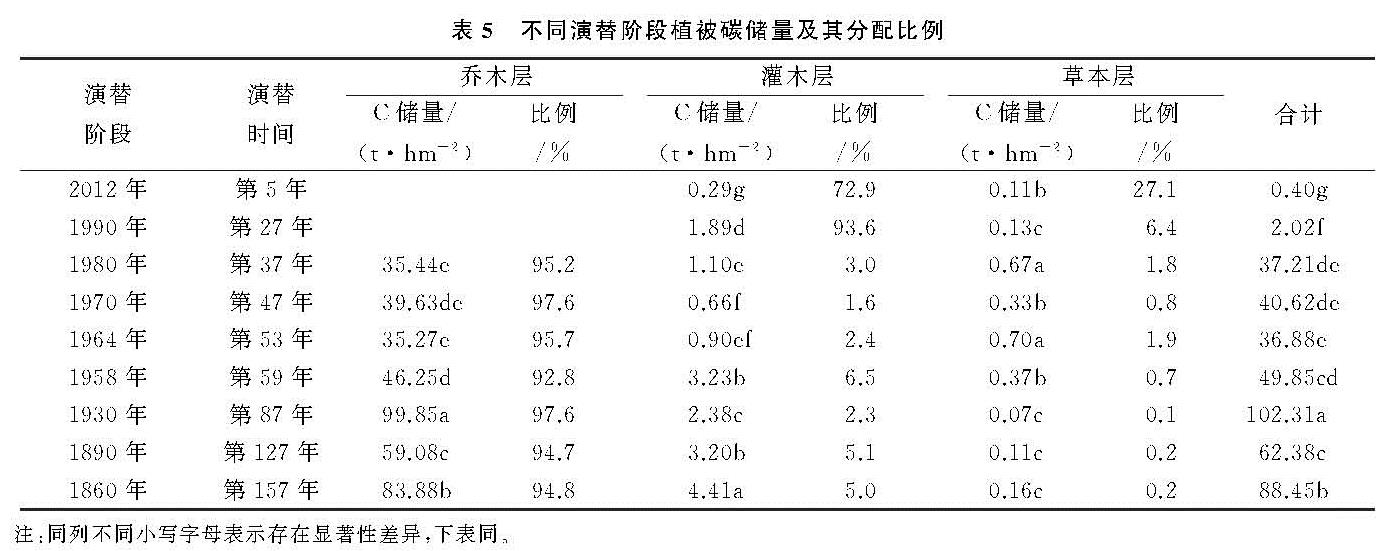
随着冰川退缩区水热条件、土壤环境的改变和植被原生演替的顺向发展,物种丰富度逐渐增大[18]。而植被生物量分配特征则主要受到其生活型和生态位的影响。本研究表明,冰川退缩裸地上的植被原生演替过程中,不同阶段的植被生物量具有显著差异。演替初期以先锋物种为主的植被生物量很低,随着演替时间的推进,植物种类增加,群落生物量总体上逐渐增加,这与杨清培等[19]对3个演替阶段上的南亚热带森林研究结果相同。然而,原生演替过程中植被生物量并不是呈线性增加,相反具有一定的波动性。本研究中植被生物量最大值出现在第87年左右,该演替阶段物种多样性最高,形成以冬瓜杨和峨嵋冷杉为主的优势种群。演替继续推进时,生物量有一定程度的下降; 说明了植被发展到一定程度时较强的自疏作用。在整个演替序列上,乔木层生物量与总生物量增长方式相同,在总生物量中占绝对优势,灌木层和草本层贡献率较低。
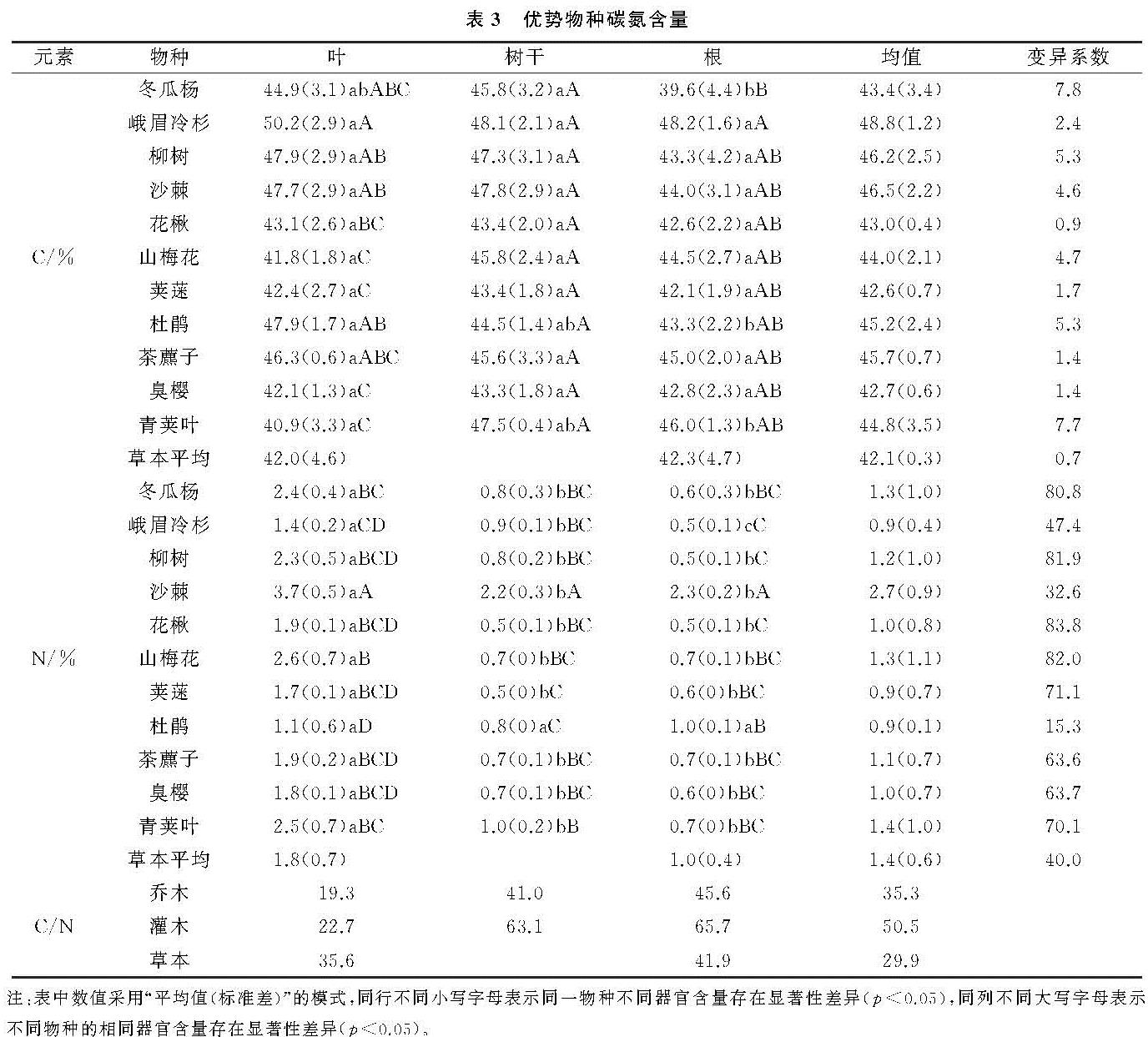
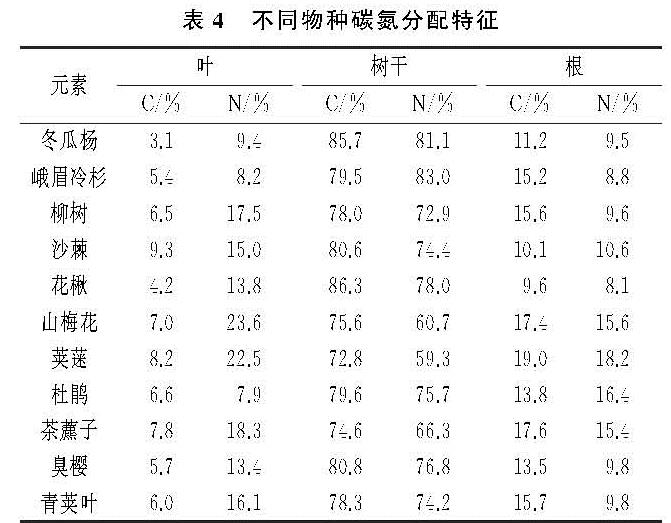
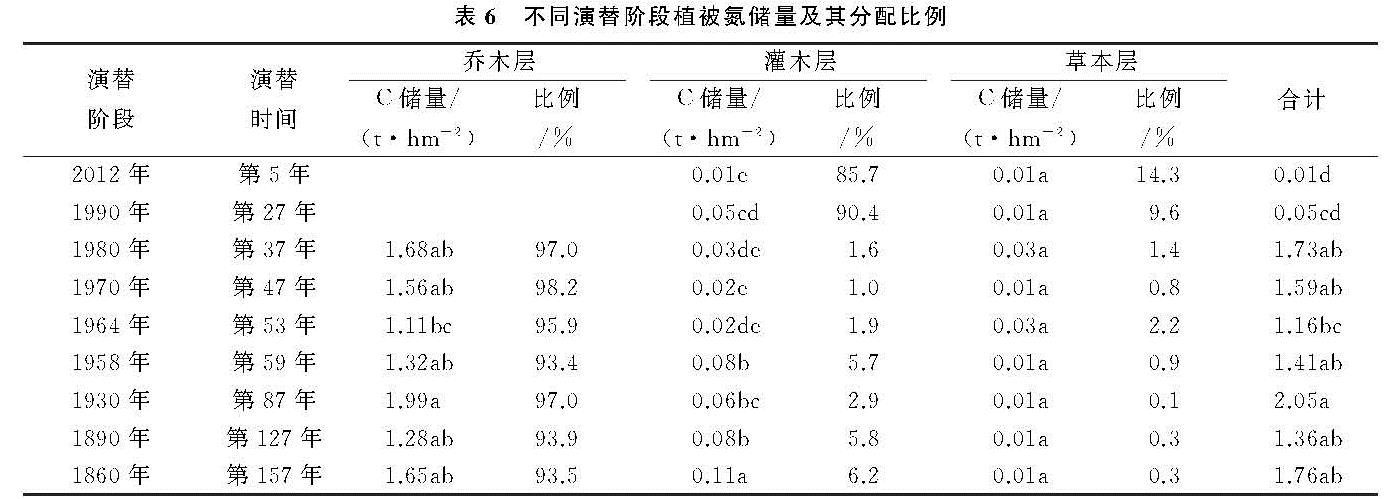
演替序列上,不同植被层、不同树种、同一树种的不同器官碳含量具有明显差异。乔木种类中,峨嵋冷杉碳含量明显高于其他物种,因为在演替中后期,峨眉冷杉较其他物种生长速度快,数量上升,相同演替阶段内积累的生物量更多,碳含量也越高。相反,沙棘氮含量较高,其原因是沙棘属于固氮植物,能进行共生固氮和氮同化[20],氮含量最高。灌木、草本对碳氮含量贡献率低。各器官碳氮含量变化无明显规律。乔木种类中,叶的碳氮含量最高,均超过生态系统植物叶的碳氮含量平均值[21-22],因为树叶通过叶绿体进行光合作用储存碳氮等有机物[23],其碳氮含量最高。根的碳氮含量最低。植物各器官的C/N中,根的C/N最大,树叶的最小,表明植被所处的生境状况良好[24],有利于演替序列上植被碳氮含量的积累。总体上,植被层C/N顺序是灌木>乔木>草本。
演替区各阶段群落碳储量的增长方式与生物量增长方式相似,呈波动性增长。其中乔木层碳氮储量最高,在各阶段活体植物碳氮储量中所占比例最大。可见,乔木层是不同演替阶段碳氮储量的主体部分,乔木层碳氮储量的增加是导致群落总碳氮随演替进行不断增加的主要原因。灌木层碳氮储量次之,草本层碳氮储量增长不明显。这与王博等[25]对库布奇东段荒漠生态系统从流动沙地到固定沙地5个不同的植被恢复演替阶段的碳氮储量的研究、杨丽韫等[26]对3个不同演替阶段的白山原始阔叶红松林碳氮储量的研究结果相一致。在整个演替序列上,由于受到森林植被生活型和生态位的影响[27],初期,草本层提供总碳氮储量。随着演替的进行,乔灌木迅速增加,碳氮储量不断增长,而草本层碳氮储量贡献率极低,呈下降趋势。一段时间后,灌木层由于资源和空间有限,开始种内种间斗争,碳氮储量有所下降。演替后期,部分乔木树种由于受气候变暖的影响,会迁移到温带或高海拔的地方[28-30],灌木数量上升,碳氮储量增加。乔木层碳氮储量一直处于上升阶段,在森林碳氮储量中占绝对优势。
研究演替序列碳氮储量动态变化,了解不同树种的相互关系,对生态系统稳定和植被恢复有一定的影响。由于海螺沟冰川退缩区对生态环境、气温与降水有很强的依赖性,群落组成复杂,缺乏对地被层、枯落物层,和土壤层的碳氮储量的研究。因此,该区森林生态系统总碳氮储量还需进一步完善。