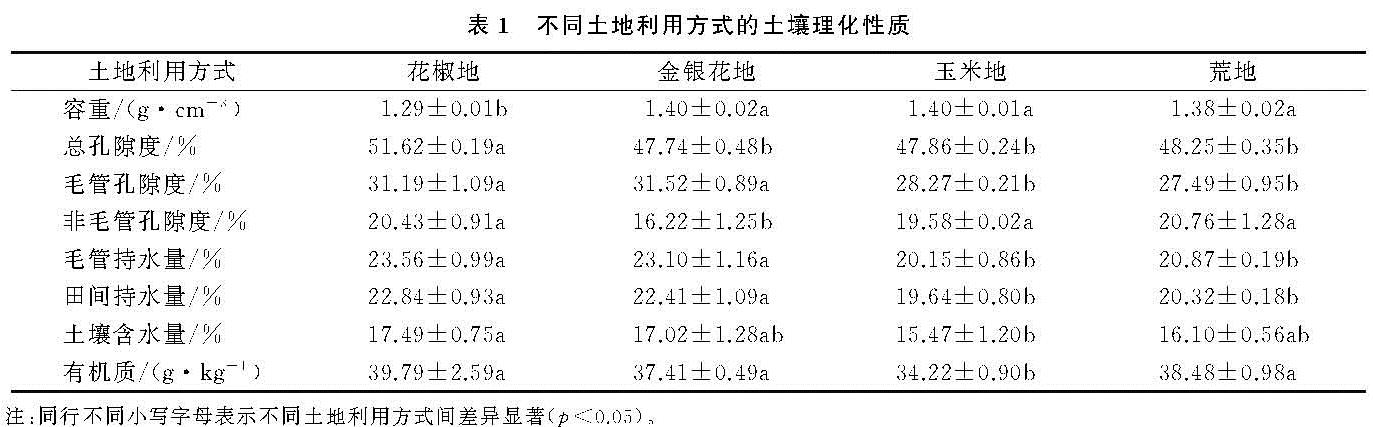
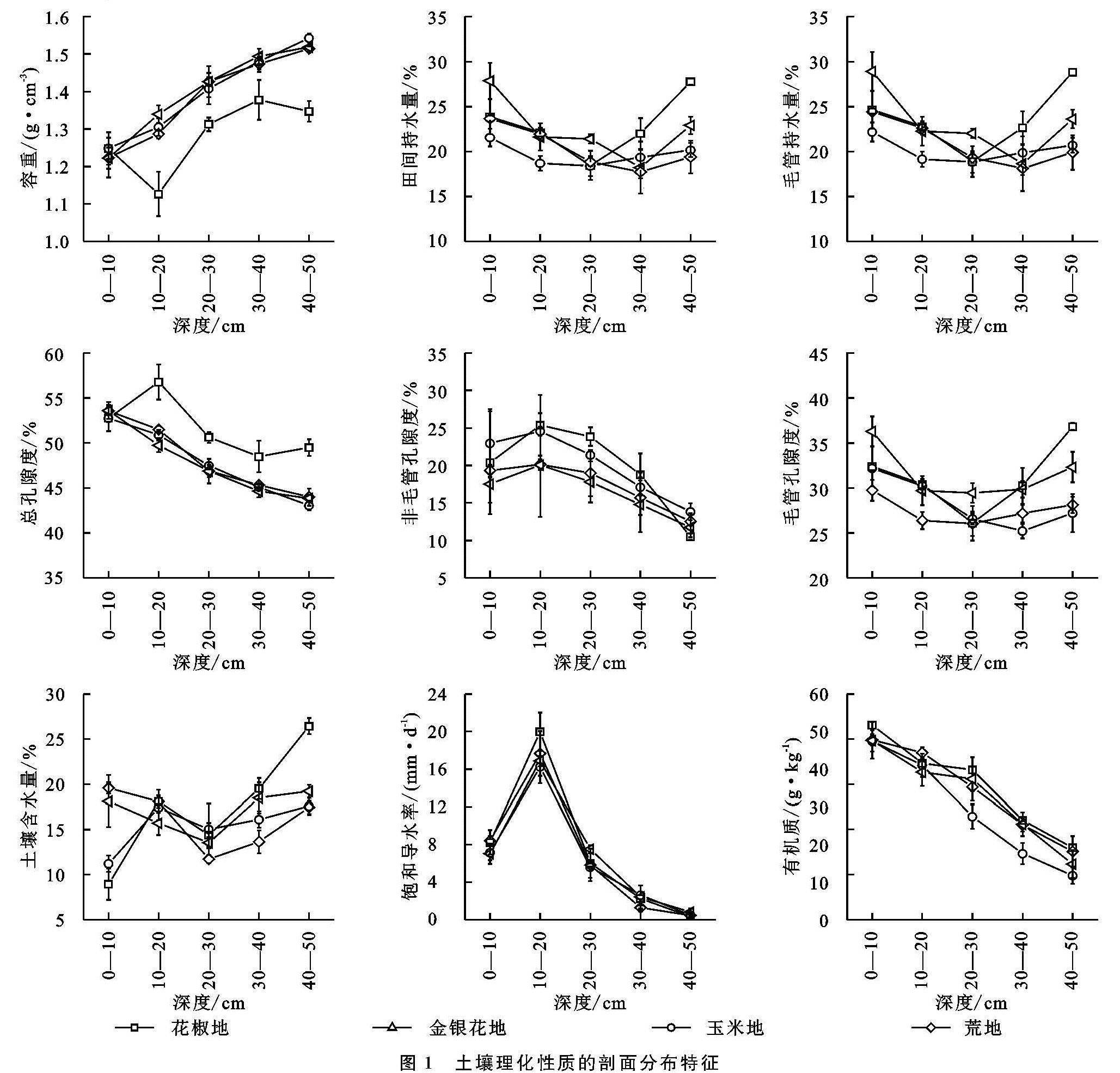
土壤Ks受到有机质、容重和孔隙度等土壤理化性质的影响。通过相关分析发现,有机质与Ks呈极显著的正相关(p<0.01)(表4),说明有机质对Ks具有正向作用。但是,关于土壤有机质对Ks的影响仍然存在争议。Lado等[24]研究认为有机质是形成和稳定土壤结构的物质,Ks随有机质含量的增加而增大。然而,可能与土壤质地差异有关,姚淑霞等[25]对科尔沁沙地研究发现Ks与有机质呈显著的负相关关系。此外,彭舜磊[8]等认为有机质提高Ks受到一定极值限制,高于极值时,Ks会呈下降趋势。尽管本研究Ks与有机质存在极显著的相关性(p<0.01),但只在10—50 cm土层,平均Ks与有机质含量变化趋势一致,主要原因是Ks受综合因素影响,可能在0—10 cm土层,非毛管孔隙度对Ks的影响起着主导的作用。
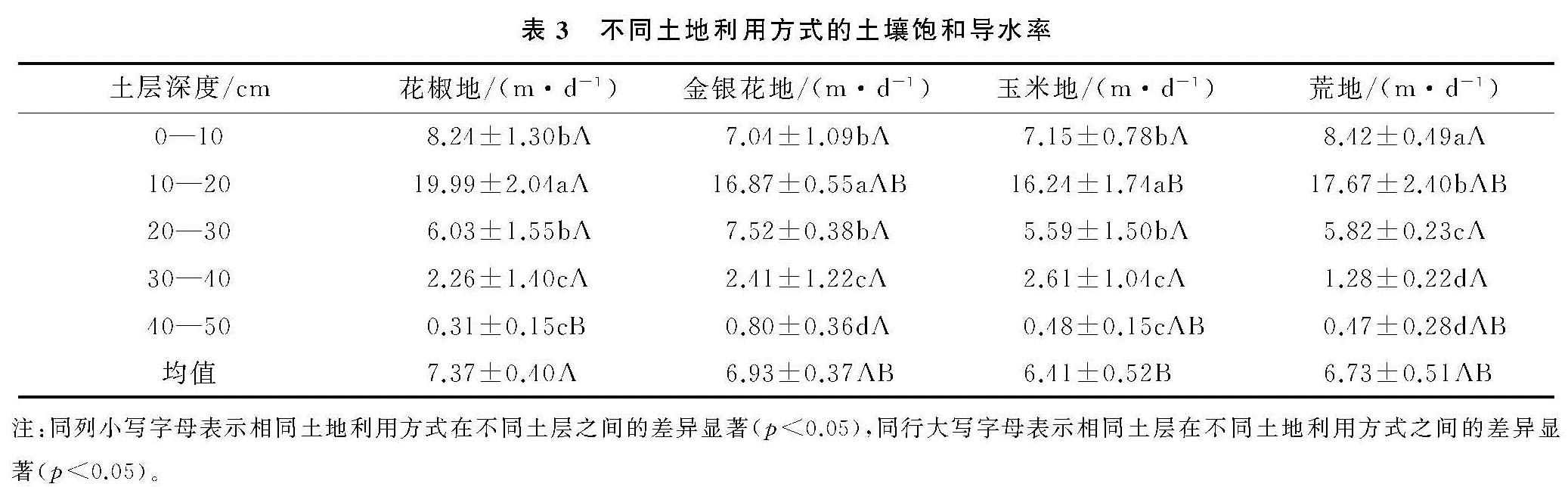
本研究Ks与容重呈极显著负相关(p<0.01),和非毛管孔隙度呈现极显著的正相关(p<0.01),这与Fu等[19]研究结果的相似。本文容重与总孔隙度、非毛管孔隙呈极显著的负相关性(p<0.01)。容重反映的是土壤孔隙特征,容重越大,反映孔隙度越小,土壤性质会通过优先传导水分的非毛管孔隙来影响Ks[6],而非毛管孔隙中的水不受束缚可以自由流动[16],利于水分的渗漏。通径分析表明,非毛管孔隙度的直接通径系数为0.719,是影响Ks最重要的因素。此外,Ks与总孔隙度也呈极显著的正相关(p<0.01)。Chen 等[26]研究表明,由于喀斯特地区土壤容重的差异,土地利用类型对土壤水力特性有较强的影响。不同土地利用方式下,植物根系发育引起的土壤质地和结构变化,是影响土壤Ks变化的重要因素[27-28]。4种土地利用方式中,土壤的Ks均随着深度的增加呈先增加后减小的变化规律(图1),平均Ks介于6.41~7.37 m/d,其中花椒地显著高于玉米地(p<0.05)(表3)。因为土壤理化性质在一定程度上受到植被的影响,植被会改变土壤结构、孔隙度而影响渗水能力。自然植被通过根系和枯枝落叶对表层土壤结构产生影响,表现在降低容重、提高有机质含量、增加大孔隙数量和增强导水能力[12]。我国西南地区树根发育主要集中在浅层土壤[29],灌木和乔木根系在地表富集,粗根向深层的横向和纵向发育,会增加非毛管孔隙度,有利于水分传导[27]。加之花椒、金银花又为浅根性植物[30-31],荒地常年杂草丛生,根系在土壤的穿插破坏土壤结构,根系死亡后形成空洞提高土壤的孔隙度。而玉米地受翻耕除草影响,植物根系因常受破坏而不发达,所以花椒地、金银花地和荒地的平均Ks高于玉米地。与其他学者研究的Ks最大值出现在表层结果不同[16],本研究各土地利用方式Ks最大值均出现在10—20 cm土层,显著高于其他土层(p<0.05)。主要原因是因为该层植被根系发达,土壤动物活动形成较多孔隙,受外界影响较小,有利于保护孔隙。但在20—50 cm土层,因土壤细颗粒向下填充,容重增大、孔隙度降低,所以Ks均随深度加深而降低。
不同土地利用方式的土壤Ks在0—10 cm土层介于7.04~8.24 m/d(表3),与付同刚等[16]研究桂北小流域的结果相近(8.72 m/d),但低于张川等[18]研究的喀斯特坡面表层(13.18 m/d)。但是在0—10 cm土层,Ks在不同土地利用方式上并未表现出显著差异(p>0.05),这是因为喀斯特地区表层土壤比深层受到更复杂的影响因素,如凋落物覆盖、动物活动[3],这些因素的综合作用会削弱表层植被对饱和导水率的影响。此外,不同土地利用方式的耕作方式差异也会对表层土壤Ks会产生重要影响。研究表明,耕作后土壤孔隙度会先下降再保持稳定[32],耕作扰动会破坏土壤结构和非毛管孔隙的连续性,特别是动物和根的生物孔[9]。本研究耕作土壤(玉米地)Ks显著低于免耕土壤(花椒地)(p<0.05),与Wang等[2]的研究结果相似。玉米地长期受到人类活动的干扰,会导致土壤层板结,有机质含量、土壤通透性降低[33]。此外,耕作扰动会加快表层细粒向下迁移,导致孔隙度随着深度加深降低,从而降低土壤Ks。综上所述,因为花椒地、金银花地和荒地均为免耕的耕作方式,植被根系发达,有机质、孔隙度高于玉米地,所以平均Ks高于玉米地。不同的研究区Ks的影响因素差异较大,主导因子不尽相同,应当根据该区的土壤理化性质具体分析。